
在哺乳动物早期发育过程中,全能性的受精卵经历第一次细胞命运决定,形成内细胞团(inner cell mass, ICM)和滋养层(trophectoderm, TE)。部分内细胞团细胞会进一步产生未来的多能性(pluripotent)细胞并最终分化出各种胚胎谱系,而滋养层则主要进一步分化为胎盘以支持胚胎发育。这两种细胞命运有各种重要的调控转录因子。体外细胞实验表明,这两类调控转录因子在促进各自命运程序的通常也会抑制对方程序。但有趣的是,在第一次细胞命运决定之前,许多ICM和TE的调控转录因子已经开始表达并且存在于同一个细胞内。那么在这个尚未分化的阶段,谱系因子是否已经开始启动各自的细胞命运程序,这些因子是否也会抑制对方的程序?早期胚胎如何解决这些因子共表达时两种细胞命运的冲突问题?由于早期胚胎材料稀缺,这些问题一直是领域内的未解之谜。
2024年1月19日,清华大学生命学院颉伟研究组在《自然结构与分子生物学》(Nature Structural & Molecular Biology)期刊上报道了题为“谱系因子TFAP2C和NR5A2在全能性胚胎中的双向激活机制”(Lineage regulators TFAP2C and NR5A2 function as bipotency activators in totipotent embryos)的研究论文,揭示了谱系转录因子在哺乳动物第一次细胞命运决定的早期起始阶段和晚期决定阶段发挥着不同的功能。该研究发现两个经典的谱系转录因子TFAP2C和NR5A2在全能性小鼠8细胞胚胎中会同时激活多能性和滋养层程序,即“双向激活”,以启动第一次细胞命运。在随后的发育过程中,谱系调控因子会逐渐转为单向激活因子,而这种转换主要是通过在基因组范围内大规模的靶基因调整来实现。
TFAP2C和NR5A2此前分别被认为属于滋养层调控因子和多能性调控因子,然而它们在具有广义全能性的8细胞胚胎中都有高表达。研究人员首先通过CUT&RUN技术捕获了包括TFAP2C和NR5A2在第一次谱系分化前后多个阶段中与染色质全基因组的结合位点。研究人员同时也构建了基因敲除(TFAP2C)以及敲低(NR5A2)的胚胎来解析这些谱系转录因子的转录调控功能。出人意料的是,研究发现在细胞命运的起始阶段,滋养层调控因子TFAP2C并没有特异性开启TE命运程序,而是同时激活了一部分表达较早的ICM和TE程序相关基因。研究人员将这种现象称为“双向激活”(bipotency activation)。
研究人员进一步探索了TFAP2C在第一次细胞命运决定后是如何促进滋养层程序的分子机制。在已分化的TE细胞中,TFAP2C脱离ICM相关基因,但保留早期TE相关基因的结合,并进一步结合和激活晚期TE相关基因,最终实现特异性激活滋养层基因。TFAP2C如何实现这一转换机制并不完全清楚,研究人员猜测可能与其他谱系特异因子的激活相关。比如CDX2是另外一个TE调控因子,其表达晚于TFAP2C。研究人员观察到TE细胞中TFAP2C和CDX2结合具有明显的共定位,同时敲除TFAP2C能够部分影响CDX2在基因组上的结合。这种转录因子之间的协助甚至可以发生在不同谱系因子之间。在植入后的胚外外胚层(ExE)中,研究者观察到TFAP2C与经典的多能性转录因子SOX2共定位并调控胚外基因的表达。TFAP2C的缺失也会影响SOX2在基因组上的结合。另外,这种谱系因子在命运决定早期“双向激活”的功能也适用于NR5A2。NR5A2在8细胞胚胎中类似地结合并激活了早期表达的ICM和TE谱系基因,而在胚胎干细胞(ESC)中则主要特异性激活ICM和多能性程序。
根据以上数据,研究人员提出,转录因子不应该被简单的定义为谱系因子。因为转录因子在不同细胞中的功能可能完全不同(比如SOX2在epiblast中能够促进胚胎谱系发育程序,但在ExE组织中能够促进胚外发育程序)。相反,多个互相影响的转录因子能够组成谱系模块(lineage module),这种模块可能是决定细胞命运的关键。单个转录因子对特定的细胞谱系的功能在谱系模块中才能被更加准确的定义。综上所述,这项研究不仅揭示了早期胚胎中独特的转录调控网络,还为转录因子在不同细胞中的功能和作用模式提供了新的视角。
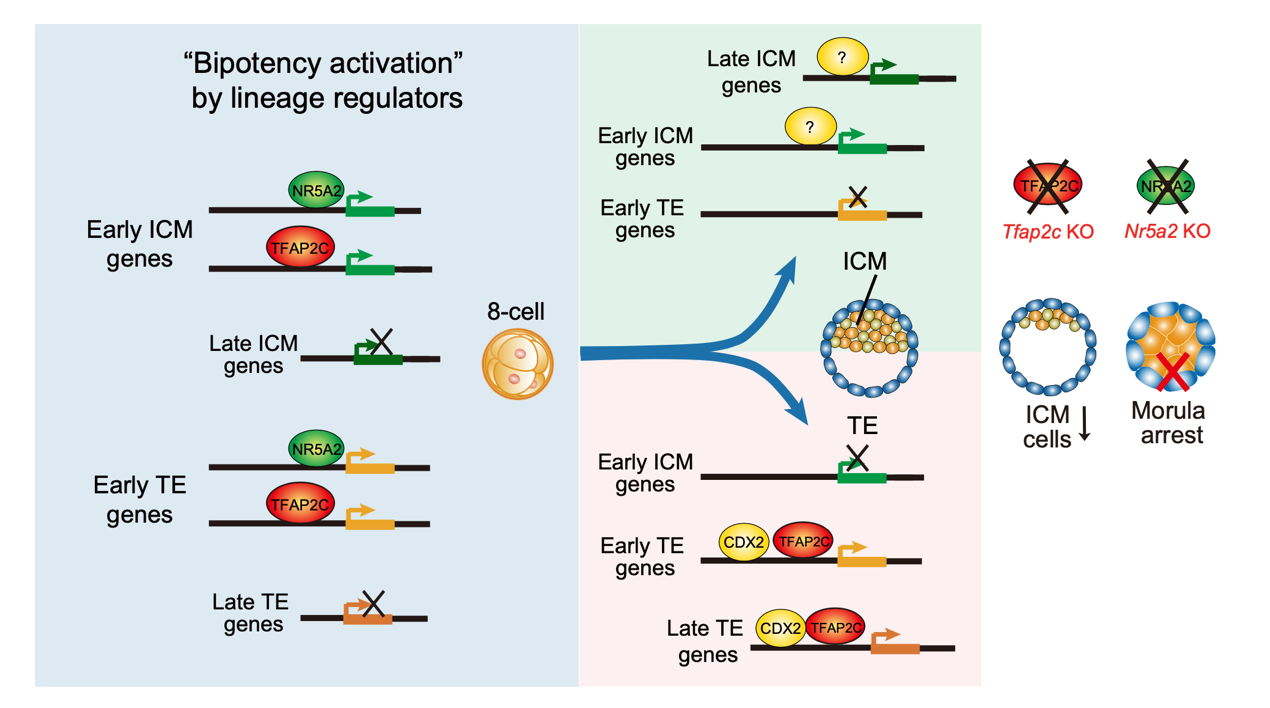
TFAP2C与 NR5A2 在早期胚胎第一次细胞命运决定前后的功能示意图
颉伟教授为本文的通讯作者,清华大学生命学院2017级博士生李丽佳(已毕业)和赖方秾(已毕业)为共同第一作者。颉伟课题组科研助理刘玲,博士后卢绪坤,2019级博士生胡晓瑜以及博士后刘伯峰也在该课题中做出重要贡献。该课题得到了清华大学实验动物中心,生物医学测试中心基因测序平台以及计算平台的大力协助和支持。本研究获得了国家自然科学基金会、国家科技部重点研发计划、清华-北京生命科学中心的经费支持。颉伟教授是HHMI国际研究学者和新基石研究员。